種類與作用
種類
核苷酸是組成核酸的基本單位,即組成核酸分子的單體。一個核苷酸分子是由一分子含氮的堿基、一分子五碳糖和一分子磷酸組成的。根據五碳糖的不同可以将核酸分為脫氧核糖核酸(DNA)和核糖核酸(RNA)兩大類。
核酸類似物
核酸類似物是與天然存在的RNA和DNA類似(結構相似)的化合物,用于醫學和分子生物學研究。核酸類似物在組成核酸的核苷酸分子以及組成核苷酸的堿基、五碳糖和磷酸基團的分子間發生了改變。通常,這些改變使得核酸類似物種的堿基配對和堿基堆積性質發生了改變。比如通用堿基可與所有四個經典堿基配對,又比如磷酸-糖骨架類似物(如PNA)甚至可形成三重螺旋。核酸類似物也稱為異種核苷酸,代表了異種生物學的主要支柱之一,即基于替代生物化學的新生自然形式的生命設計。
核酸類似物包括肽核酸(PNA),嗎啉代和鎖核酸(LNA)以及乙二醇核酸(GNA)和蘇糖核酸(TNA)。因為分子主鍊發生了改變,它們與天然存在的DNA或RNA有明顯的不同。
作用
DNA是儲存、複制和傳遞遺傳信息的主要物質基礎。
RNA在蛋白質合成過程中起着重要作用——其中轉運核糖核酸,簡稱tRNA,起着攜帶和轉移活化氨基酸的作用;信使核糖核酸,簡稱mRNA,是合成蛋白質的模闆;核糖體的核糖核酸,簡稱rRNA,是細胞合成蛋白質的主要場所。
此外,現在已知許多其他種類的功能RNA,如microRNA等。
核酸類似物主要用于醫學和分子生物學研究。
曆史
核酸的發現
核酸最早于1869年由瑞士醫生和生物學家弗雷德裡希·米歇爾分離獲得,稱為Nuclein。
在19世紀80年代早期,德國生物化學學家,1910年諾貝爾生理和醫學獎獲得者科塞爾進一步純化獲得核酸,發現了它的強酸性。他後來也确定了核堿基。
1889年,德國病理學家Richard Altmann創造了核酸這一術語,取代了Nuclein。
1919年,一位美籍俄羅斯醫生和化學家菲巴斯·利文首先發現了單核苷酸的三個主要成分(磷酸鹽、戊糖和氮基)的順序。
1938年,英國物理學家和生物學家威廉·阿斯特伯裡和Florence Bell(後來改名為Florence Sawyer)發表了第一個DNA的X射線衍射圖譜。
1953年,美國分子生物學家詹姆斯·沃森和英國分子生物學家弗朗西斯·克裡克确定了DNA的結構。
核酸的實驗研究構成了現代生物學和醫學研究的重要組成部分,并為基因組和法醫學以及生物技術和制藥工業奠定了基礎。
分子大小及組成
分子大小
核酸分子通常很大。實際上,DNA分子可能是已知的最大的單個生物分子。
但也有比較小的核酸分子。
核酸分子的大小範圍從21個核苷酸(小幹擾RNA)到大染色體(人類染色體是一個含有2.47億個堿基對的單個分子)不等。
化學組成
核酸完全水解産生嘌呤和嘧啶等堿性物質、戊糖(核糖或脫氧核糖)和磷酸的混合物。核酸部分水解則産生核酸和核苷酸。每個核苷分子含一分子堿基和一分子戊糖,一分子核苷酸部分水解後除産生核苷外,還有一分子磷酸。
DNA和RNA含有的核糖同,DNA含有脫氧核糖,而RNA含有核糖。此外,DNA和RNA中含有的堿基也有差别:DNA和RNA都含有腺嘌呤,胞嘧啶和鳥嘌呤,但DNA中不含有尿嘧啶,隻有胸腺嘧啶
核酸中的糖和磷酸鹽通過磷酸二酯鍵以交替鍊(糖-磷酸骨架)相互連接。磷酸基團所連接的碳是糖的3'-末端,與碳原子結合的碳是5'-末端,這就産生了核酸的方向性。核堿基通過N-糖苷鍵與糖連接。
在RNA和DNA中也發現了非标準核苷,它們通常來自DNA分子内的标準核苷或初始RNA轉錄物的修飾。轉移RNA(tRNA)分子含有特别多的修飾核苷。
分子組成
天然存在的DNA分子在大多數情況下是雙鍊的,而RNA分子是單鍊的。然而,有許多例外。一些病毒具有由雙鍊RNA構成的基因組,而其他病毒具有單鍊DNA基因組,并且在某些情況下,可形成具有三個或四個鍊的核酸結構。
相關性質
化學性質
酸效應:在強酸和高溫下核酸完全水解為堿基,核糖或脫氧核糖和磷酸。在濃度略稀的無機酸中,最易水解的化學鍵被選擇性的斷裂,一般為連接嘌呤和核糖的糖苷鍵,從而産生脫嘌呤核酸。
堿效應:當pH值超出生理範圍(pH7~8)時,對DNA結構将産生更為微妙的影響。堿效應使堿基的互變異構态發生變化。這種變化影響到特定堿基間的氫鍵作用,結果導緻DNA雙鍊的解離,稱為DNA的變性。pH較高時,同樣的變性發生在RNA的螺旋區域中,但通常被RNA的堿性水解所掩蓋。
化學變性:一些化學物質能夠使DNA或RNA在中性pH下變性。由堆積的疏水堿基形成的核酸二級結構在能量上的穩定性被削弱,則核酸變性。
物理性質
黏性:DNA的高軸比等性質使得其水溶液具有高黏性,很長的DNA分子又易于被機械力或超聲波損傷,同時黏度下降。
浮力密度:可根據DNA的密度對其進行純化和分析。在高濃度分子質量的鹽溶液(CsCl)中,DNA具有與溶液大緻相同的密度,将溶液高速離心,則CsCl趨于沉降于底部,從而建立密度梯度,而DNA最終沉降于其浮力密度相應的位置,形成狹帶,這種技術成為平衡密度梯度離心或等密度梯度離心。
穩定性:核酸的結構相當穩定,其主要原因有堿基對間的氫鍵、堿基的堆積作用和環境中的陽離子。
光譜學性質
減色性:dsDNA相對于ssDNA是減色的,而ssDNA相對于dsDNA是增色的。
DNA純度:通過測量A260/A280和A260/A230進行判斷。
變性、複性和雜交
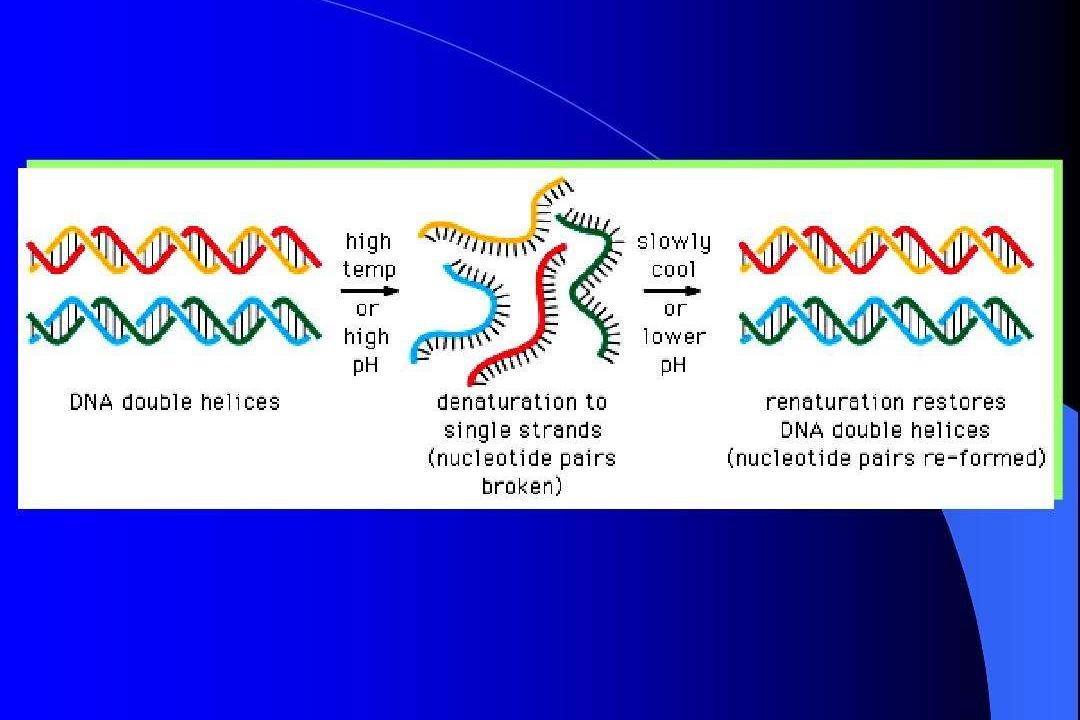
變性
在一定理化因素作用下,核酸雙螺旋等空間結構中堿基之間的氫鍵斷裂,變成單鍊的現象稱為變性(denaturation)。
引起核酸變性的常見理化因素有加熱、酸、堿、尿素和甲酰胺等。在變性過程中,核酸的空間構象被破壞,理化性質發生改變。由于雙螺旋分子内部的堿基暴露,其A260值會大大增加。A260值的增加與解鍊程度有一定比例關系,這種關系稱為增色效應(hyperchromic effect)。如果緩慢加熱DNA溶液,并在不同溫度測定其A260值,可得到“S”形DNA熔化曲線(melting curve)。從DNA熔化曲線可見DNA變性作用是在一個相當窄的溫度内完成的。
當A260值開始上升前DNA是雙螺旋結構,在上升區域分子中的部分堿基對開始斷裂,其數值随溫度的升高而增加,在上部平坦的初始部分尚有少量堿基對使兩條鍊還結合在一起,這種狀态一直維持到臨界溫度,此時DNA分子最後一個堿基對斷開,兩條互補鍊徹底分離。通常把加熱變性時DNA溶液A260升高達到最大值一半時的溫度稱為該DNA的熔解溫度(melting temperature Tm),Tm是研究核酸變性很有用的參數。Tm一般在85~95℃之間,Tm值與DNA分子中GC含量成正比。
複性
變性DNA在适當條件下,可使兩條分開的單鍊重新形成雙螺旋DNA的過程稱為複性(renaturation)。當熱變性的DNA經緩慢冷卻後複性稱為退火(annealing)。DNA複性是非常複雜的過程,影響DNA複性速度的因素很多:DNA濃度高,複性快;DNA分子大複性慢;高溫會使DNA變性,而溫度過低可使誤配對不能分離等等。最佳的複性溫度為Tm減去25℃,一般在60℃左右。離子強度一般在0.4mol/L以上。
雜交
具有互補序列的不同來源的單鍊核酸分子,按堿基配對原則結合在一起稱為核酸雜交(hybridization)。雜交可發生在DNA-DNA、RNA-RNA和DNA-RNA之間。雜交是分子生物學研究中常用的技術之一,利用它可以分析基因組織的結構,定位和基因表達等,常用的雜交方法有Southern印迹法,Northern印迹法和原位雜交等。