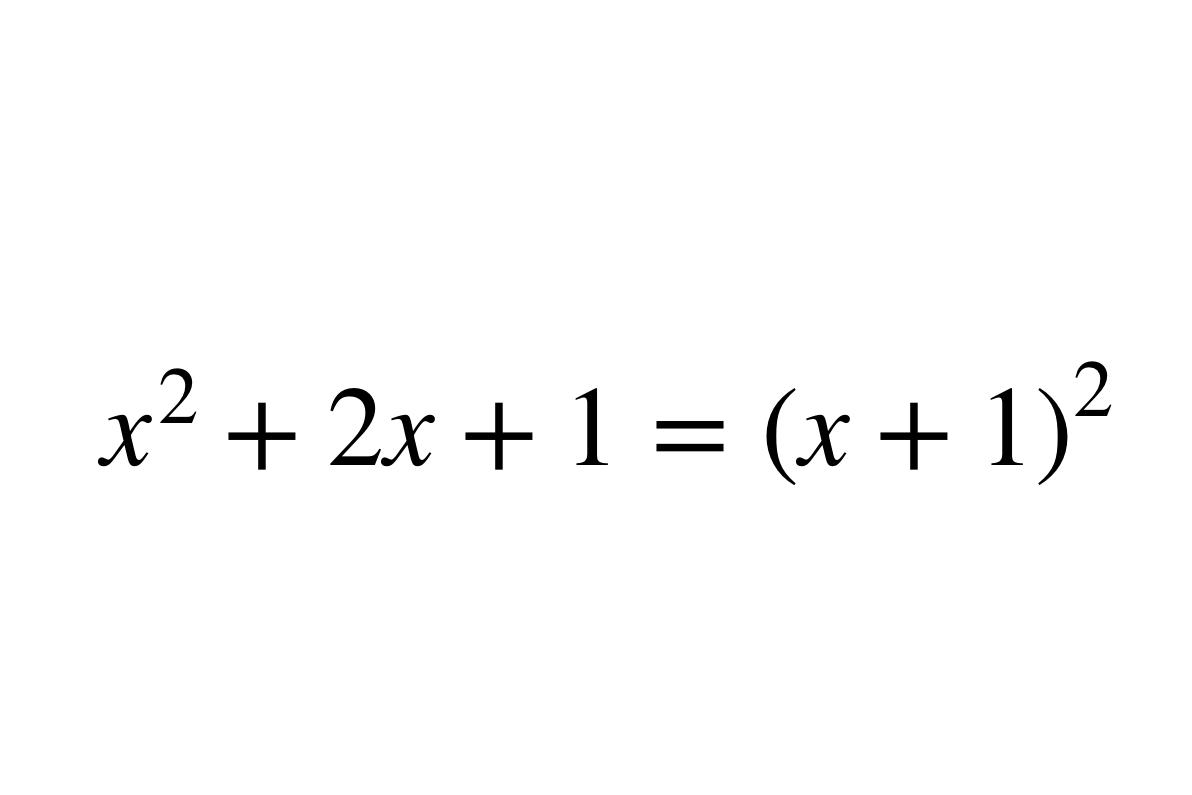
概述
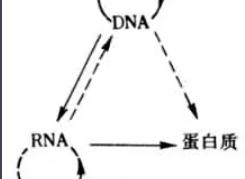
基因是能够自我复制,永远保存的单位,它的生理功能是以蛋白质的形式表达出来的。所以DNA核苷酸序列是遗传信息的储存者,它通过自主复制得以永存,通过转录生成信使RNA,进而翻译成蛋白质的过程来控制生命现象,即贮存在核酸中的遗传信息通过转录,翻译成为蛋白质,体现为丰富多彩的生物界,这就是生物学中的中心法则(central dogma)。该法则表明信息流的方向是有DNA→RNA→蛋白质。在该信息流中,RNA病毒及某些动物细胞可以RNA为模板复制出RNA,然后再由RNA直接合成出蛋白质;同时某些病毒,某些癌细胞及动物胚胎细胞可以由RNA转录出DNA,即发生反转录(reverse transcription)。由中心法则可知DNA(基因)控制着蛋白质的合成。
介绍
遗传信息在细胞内的生物大分子间转移的基本法则。包含在脱氧核糖核酸(DNA)或核糖核酸(RNA)分子中的具有功能意义的核苷酸顺序称为遗传信息。遗传信息的转移包括核酸分子间的转移、核酸和蛋白质分子间的转移。
早在1909年,伽罗德(A•E•Garrod)在《先天性代谢差错》一书中,就描述了黑尿病基因与尿黑酸氧化酶的关系。以红色面包霉(链孢霉)为材料而开创生化遗传学研究的比德尔(G•W•Beadle),1941年与塔特姆(E•L•Tatum)一起提出“一个基因一种酶”的假说,认为基因是通过酶来起作用的。 基因(DNA)主要位于细胞核中。如果酶(化学本质是蛋白质)是在细胞核内合成的,问题倒也简单,由基因直接指导酶的合成就是了。可事实却并不如此。
早在40年代,汉墨林(J•Hammerling)和布拉舍(J•Brachet)就分别发现伞藻和海胆卵细胞在除去细胞核之后,仍然能进行一段时间的蛋白质合成。这说明细胞质能进行蛋白质合成。
1955年李托菲尔德(Littlefield)和1959年麦克奎化(K•McQuillen)分别用小鼠和大肠杆菌为材料证明细胞质中的核糖体是蛋白质合成的场所。这样,细胞核内的DNA就必须通过一个“信使”(message)将遗传信息传递到细胞质中去。
1955年,布拉舍用洋葱根尖和变形虫为材料进行实验,他用核糖核酸酶(RNA酶)分解细胞中的核糖核酸(RNA),蛋白质的合成就停止。而如果再加入从酵母中抽提的RNA,蛋白质的合成就有一定程度的恢复。同年,戈尔德斯坦(Goldstein)和普劳特(Plaut)观察到用放射性标记的RNA从细胞核转移到细胞质。因此,人们猜测RNA是DNA与蛋白质合成之间的信使。1961年,雅可布(F•Jacob)和莫诺(J•Monod)正式提出“信使核糖核酸”(mRNA)的术语和概念。
1964年马贝克斯(C•Marbaix)从兔的网织红细胞中分离出一种分子量较大而寿命很短的RNA,被认为是mRNA。实际上,早在1947年,法国科学家布瓦旺(A•Boivin)和旺德雷利(R•Vendrely)就在当年的《实验》杂志上联名发表了一篇论文,讨论DNA、RNA与蛋白质之间可能的信息传递关系。一位不知名的编辑把这篇论文的中心思想理解为DNA制造了RNA,再由RNA制造蛋白质。
10年以后,1957年9月,克里克提交给实验生物学会一篇题为“论蛋白质合成”的论文,发表在该学会的论文集(Symposum of the Society for Experimental Biology)第12卷第138页。这篇论文被评价为“遗传学领域最有启发性、思想最解放的论著之一。”在这篇论文中,克里克正式提出遗传信息流的传递方向是DNA→RNA→蛋白质,后来被学者们称为“中心法则”。
1957年F.H.C.克里克最初提出的中心法则是:DNA→RNA→蛋白质它说明遗传信息在不同的大分子之间的转移都是单向的,不可逆的,只能从DNA到RNA(转录),从RNA到蛋白质(翻译)。这两种形式的信息转移在所有生物的细胞中都得到了证实。
生物遗传中心法则最早是由Crick于1958年提出的,用以表示生命遗传信息的流动方向或传递规律。由于当时对转录、翻译、遗传密码、肽链折叠等都还了解不多,在那个时候中心法则带有一定的假设性质。随着生物遗传规律的进一步探索,中心法则也逐步得到完善和证实。
1970年H.M.特明和D.巴尔的摩在一些RNA致癌病毒中发现它们在宿主细胞中的复制过程是先以病毒的RNA分子为模板合成一个DNA分子,再以DNA分子为模板合成新的病毒RNA。前一个步骤被称为反向转录,是上述中心法则提出后的新的发现。因此克里克在1970年重申了中心法则的重要性,提出了更为完整的图解形式。
这里遗传信息的转移可以分为两类:第一类用实线箭头表示,包括DNA的复制、RNA的转录和蛋白质的翻译,即①DNA→DNA(复制);②DNA→RNA(转录);③RNA→蛋白质(翻译)。这三种遗传信息的转移方向普遍地存在于所有生物细胞中。第二类用虚线箭头表示,是特殊情况下的遗传信息转移,包括RNA的复制,RNA反向转录为DNA和从DNA直接翻译为蛋白质。即①RNA→RNA(复制);②RNA→DNA(反向转录);③DNA→蛋白质。RNA复制只在RNA病毒中存在。反向转录最初在RNA致癌病毒中发现,现在在人的白细胞和胎盘滋养层中也测出了与反向转录有关的反向转录酶的活性。至于遗传信息从DNA到蛋白质的直接转移仅在理论上具可能性,在活细胞中尚未发现。遗传信息在细胞内生物大分子之间转移的基本法则。认为遗传信息的转移只能从脱氧核糖核酸到核糖核酸(转录),从核糖核酸到蛋白质(转译)。现已发现,在某些生物中,核糖核酸通过逆转录,也能把遗传信息传递给脱氧核糖核酸。
人们知道,细胞可以将DNA复制成一组新的DNA,然后进入一个新形成的细胞。其中,涉及一类被称为聚合酶的细胞“机器”,它们也可以构建RNA信息,这些从DNA中心库复制的信息可以被更有效地“解读”为蛋白质。但聚合酶被认为只沿着DNA到DNA或DNA到RNA的方向起作用,从而防止RNA信息被重写回DNA中心库中。现在,研究人员首次证明RNA也可以被写回DNA,这挑战了生物学的核心法则。
发展史
①1965年,科学家发现RNA可复制;
②1970年,科学家发现逆转录酶;
③1982年,科学家发现疯牛病是由一种结构异常的蛋白质引起的疾病。
内容
①从DNA流向DNA(DNA自我复制);
②从DNA流向RNA,进而流向蛋白质(转录和翻译);
③从RNA流向RNA(RNA自我复制);
④从RNA流向DNA(逆转录);
注:其中前两条是中心法则的主要体现,后两条是中心法则的完善和补充。
朊粒
朊粒是一种蛋白质传染颗粒(proteinaceous infectious particle),它最初被认识到是羊的瘙痒病的病原体。这是一种慢性神经系统疾病,在200多年前就已发现。1935年法国研究人员通过接种发现这种病可在羊群中传染,意味着这种病原体是能在宿主动物体内自行复制的感染因子。朊粒同时又是人类的中枢神经系统退化性疾病如库鲁病(Kuru)和克—杰氏综合征(Creutzfeldt-Jacobdisease,CJD)的病原体,也可引起疯牛病即牛脑的海绵状病变(bovin spongiform encephalopathy,BSE)。以后的研究证明,这种朊粒不是病毒,而是不含核酸的蛋白质颗粒。一个不含DNA或RNA的蛋白质分子能在受感染的宿主细胞内产生与自身相同的分子,且实现相同的生物学功能,即引起相同的疾病,这意味着这种蛋白质分子也是负载和传递遗传信息的物质。这是从根本上动摇了遗传学的基础。
实验证明,朊粒确实是不含DNA和RNA的蛋白质颗粒,但它不是传递遗传信息的载体,也不能自我复制,而仍是由基因编码产生的一种正常蛋白质的异构体。
基因编码
哺乳动物细胞里的基因编码产生一种糖蛋白PrP。人的PrP基因位于20号染色体短臂,PrP由253个氨基酸残基组成,在氨基端有22个氨基酸组成的信号肽。在正常脑组织中的PrP称为PrPc,相对分子质量为33000~35000,对蛋白酶敏感。在病变脑组织中的PrP称为PrPsc,相对分子质量为27000~30000,是PrPc中的一段,蛋白酶对其不起作用。现在知道,PrPc和PrPsc是PrP的两种异构体,氨基酸组分和线性排列次序相同,但是三维构象不同。PrPc的结构中。螺旋占42%,β片层占30%;PrPsc则是。螺旋占30%,β片层占43%。PrPc的4条。螺旋可以排列成一个致密的球状结构,这个结构的随机涨落(stochastic fluctua—tion)会长成部分折叠的单体PrP*,这是一种中间体,即PrP*可以生成PrPc,也可以生成PrPsc。一般情况下,PrP*的含量极少,所以生成的PrPsc极少。可是外源的PrPsc可以促使PrP*变成PrPsc。PrPsc的不溶性使生成PrPsc过程成为不可逆转。PrPsc在神经细胞里大量沉积,引起神经细胞的病变,破坏了神经细胞功能。因此,PrPsc感染正常细胞后,可以促使细胞内生成更多的PrPsc,PrPsc逐渐积累,需要有一个时间过程才会引发疾病,这也就是这种神经退化性疾病有一个很长的潜伏期的原因。所以说,PrPsc进入宿主细胞并不是自我复制,而是将细胞内基因编码产生的PrPc变成PrPsc。因此,中心法则是正确的,至少在目前还是无需修正的。
基因表达
关系
基因指导蛋白质合成;基因控制生物体;生物体性状由蛋白质直接体现。
调控方法
a.基因通过控制酶的合成来控制代谢过程,进而控制生物体性状。
b.基因通过指导蛋白质的合成,控制蛋白质结构进而直接控制生物体的性状。
意义
中心法则是现代生物学中最重要最基本的规律之一,其在探索生命现象的本质及普遍规律方面起了巨大的作用,极大地推动了现代生物学的发展,是现代生物学的理论基石,并为生物学基础理论的统一指明了方向,在生物科学发展过程中占有重要地位。遗传物质可以是DNA,也可以是RNA。细胞的遗传物质都是DNA,只有一些病毒的遗传物质是RNA。这种以RNA为遗传物质的病毒称为反转录病毒(retrovirus),在这种病毒的感染周期中,单链的RNA分子在反转录酶(reverse transcriptase)的作用下,可以反转录成单链的DNA,然后再以单链的DNA为模板生成双链DNA。双链DNA可以成为宿主细胞基因组的一部分,并同宿主细胞的基因组一起传递给子细胞。在反转录酶催化下,RNA分子产生与其序列互补的DNA分子,这种DNA分子称为互补DNA(complementary DNA),简写为cDNA,这个过程即为反转录(reverse transcription)。
任何一种假设都要经受科学事实的检验。反转录酶的发现,使中心法则对关于遗传信息从DNA单向流入RNA做了修改,遗传信息是可以在DNA与RNA之间相互流动的。那么,对于DNA和RNA与蛋白质分子之间的信息流向是否只有核酸向蛋白质分子的单向流动,还是蛋白质分子的信息也可以流向核酸,中心法则仍然肯定前者。可是,病原体朊粒(Prion)的行为曾对中心法则提出了严重的挑战。
由此可见,遗传信息并不一定是从DNA单向地流向RNA,RNA携带的遗传信息同样也可以流向DNA。但是DNA和RNA中包含的遗传信息只是单向地流向蛋白质,迄今为止还没有发现蛋白质的信息逆向地流向核酸。这种遗传信息的流向,就是克里克概括的中心法则(central dogma)的遗传学意义。
遗传信息的特殊传递
逆转录
主条目:逆转录
在中心法则被详细阐述之后,人们发现了逆转录病毒。这些病毒可通过一种叫做逆转录酶的催化,以RNA为模板逆转录合成cDNA再由cDNA转录出RNA。这肯定了RNA向DNA转录的存在。人们最初以为这种现象仅出现于病毒中,但在最近,在高等动物中亦发现了RNA向DNA转录的逆转录转座子。
RNA复制
主条目:RNA复制
有些病毒的遗传物质是RNA分子,靠RNA复制而传代,以RNA为模板的RNA复制酶催化下合成RNA分子,RNA复制酶中缺乏校正功能,复制时错误率很高。RNA复制酶只对病毒本身的RNA起作用,而不会作用于宿主细胞中的RNA分子。
RNA的催化功能
主条目:RNA催化
人们一直认为生物体内的各种生化反应都是由酶来催化完成的,而RNA仅是存贮与传递信息,与酶的催化反应无关。核糖核酸酶P是一种核酶,即由一个RNA分子发挥催化活性,它是第一个被发现的蛋白质以外具有催化活性的生物大分子。它的功能是剪切tRNA分子中RNA上多余的或前体的多余序列。RNA可以不通过蛋白质而直接表现出本身的某些遗传信息,而这种信息并不是以核苷酸三联密码来编码。
直接以DNA为模板合成蛋白质
有人在一些离体实验中观察到,一些与蛋白质合成抑制剂类抗生素如新霉素和链霉素,能扰乱核糖体对信使的选择,从而可以接受单链DNA分子代替mRNA,然后以单链DNA为模版,按核苷酸顺序转译成多肽的氨基酸顺序。另外还有研究表明,细胞核里的DNA可以直接转移到细胞质中的核糖体上,不需要通过RNA也可以控制蛋白质的合成。
DNA也具有酶活性
1994年乔依斯(G.F.Joyce)等人发现一个人工合成的DNA分子具有一种特殊的磷酸二酯酶活性。此后又有多例报道人工合成的DNA序列具有各种不同的酶活性。1995年中国学者王身立等人发现从多种生物中提取的DNA均具有酯酶活性,能催化乙酸萘酯水解为萘酚和乙酸。这种较弱的酯酶活性是非特异性DNA的一般性质,并不需要特定序列的DNA编码。
中心法则的扩充
克里克在上述那篇1970年的文章中指出,中心法则虽然对指导实验很有用,但不应该被当成教条:
“虽然本文所提出的各类法则看来是可靠的,可是我们对分子生物学的认识,即使只是一个细胞—更不用说大自然里的整个生命体—仍然远远未完备到,足以让我们把它当成教条一样肯定正确的程度”
——克里克
自从克里克发表1970年那篇文章以来,很多新发现说明了中心法则补充和发展的必要。
翻译后修饰
主条目:翻译后修饰
对于大部份的蛋白质来说,这是蛋白质生物合成的最后步骤。蛋白质的翻译后修饰会附上其他的生物化学官能团、改变氨基酸的化学性质,或是造成结构的改变来扩阔蛋白质的功能。酶可以从蛋白质的N末端移除氨基酸,或从中间将肽链剪开。举例来说,胰岛素是肽的激素,它会在建立双硫键后被剪开两次,并在链的中间移走多肽前体,而形成的蛋白质包含了两条以双硫键连接的多肽链。其他修饰,就像磷酸化,是控制蛋白质活动机制的一部份。蛋白质活动可以是令酶活性化或钝化。
蛋白质的内含子
主条目:蛋白质内含子
蛋白质有自剪接现象,与mRNA相同,一些蛋白质前体具有内含子(intein)序列,多肽序列中间的某些区域被加工切除,剩余部分的蛋白质外显子(extein)重新连接为蛋白质分子。
DNA甲基化
主条目:表观遗传学
表观遗传学研究在没有细胞核DNA序列改变的情况时,基因功能的可逆的、可遗传的改变。这些改变包括DNA的修饰(如甲基化修饰)、RNA干扰、组蛋白的各种修饰等。也指生物发育过程中包含的程序的研究。在这两种情况下,研究的对象都包括在DNA序列中未包含的基因调控信息如何传递到(细胞或生物体的)下一代这个问题。其主要研究内容包括大致两方面内容。一类为基因选择性转录表达的调控,有DNA甲基化,基因印记,组蛋白共价修饰,染色质重塑。另一类为基因转录后的调控,包含基因组中非编码的RNA,微小RNA,反义RNA,内含子及核糖开关等。
主条目:DNA甲基化
DNA甲基化为DNA化学修饰的一种形式,能在不改变DNA序列的前提下,改变遗传表现。为外遗传编码(epigenetic code)的一部分,是一种外遗传机制。DNA甲基化过程会使甲基添加到DNA分子上,例如在胞嘧啶环的5'碳上:这种5'方向的DNA甲基化方式可见于所有脊椎动物。
蛋白质可作为合成DNA的模板
来自Mount.Sinai医院的研究人员发现了一种叫Rev1 DNA聚合酶的蛋白质,它可以为DNA复制提供编码信息。许多致癌物质会倾向于破坏DNA的鸟嘌呤(G),或者是破坏鸟嘌呤与胞嘧啶(C)的配对,这些都会导致DNA错配的发生。新发现的蛋白质可以以自身为模板在复制链上加一个胞嘧啶,这个胞嘧啶无论鸟嘌呤是否在DNA链中存在都会被Rev1加上去的,在DNA复制时可以利用一条单链,根据碱基配对原则复制出新的DNA链。细胞利用这种崭新的机制在含有致癌物质的情况下对受损的DNA进行复制。这是第一次发现蛋白质可以作为一种合成DNA的模板。
朊病毒
主条目:朊病毒
朊病毒是通过改变其他蛋白质的构象来进行自身精确复制的一类蛋白质。也就是:蛋白质→蛋白质。这种具有感染性的因子主要由蛋白质组成。具有感染性的因子Prp与正常因子PrP在形状上有一点不同。科学家推测这种变形的蛋白质会引起正常的PrP转变成具有感染性的蛋白质,这种连锁反应使得正常的蛋白质和致病的蛋白质因子都成为新病毒。
中心法则的起源
中心法则的信息是从DNA到RNA,但是,谢平(2014)指出,从生命起源和演化的历史来看,信息的整合则必定是从mRNA到DNA 。
从RNA到DNA的演化之路
在细胞起源的早期,为了适应细胞的分裂行为,遗传物质的有效传递成为必须,因此,细胞中储存在各种m-RNA中的遗传信息的整合必须成为选择的方向,把所有m-RNA的信息连接起来,就是向DNA方向发展的启航。也许可以认为,随着蛋白质的增多,mRNA也相应增多,偶尔一个整合性的mRNA长链更好地匹配了细胞的分裂行为,这样就会得到选择。
但是,并不是把m-RNA拼接起来就是DNA,实际上,结构成份发生了两个变化,其一,RNA分子中的尿嘧啶,在DNA中变成了胸腺嘧啶,虽然两者仅有细微的差别,即后者多了一个甲基;其二,RNA分子中的核糖在DNA中变成了脱氧核糖。但是这两个变化却导致了两种核酸在形态上的显著差别:DNA形成双螺旋的结构,而绝大部分RNA分子都是线状单链,虽然RNA分子的某些区域可自身回折进行碱基互补配对,形成局部双螺旋。或许出于某种结构上的缘由,如果脱氧核糖替代核糖以及胸腺嘧啶替代尿嘧啶能更加有利于形成稳定的双螺旋结构的话,那就是DNA被选择的方向性。
当然,或许仅仅就是为了避免混淆,因为生物经常要用既有联系又能区别的结构物质来行使不同的功能,譬如,NADPH和NADH,两者的还原电位完全相同,功能也类似,但却用于不同的生物代谢途径。一个不容易混淆的井然有序的代谢系统当然会得到选择或青睐 。
用mRNA拼接DNA的证据
真核基因转录产物为单顺反子,即一个基因编码一条多肽链或RNA链,每个基因转录有各自的调节元件。在原核细胞中,通常是几种不同的mRNA连在一起,相互之间由一段短的不编码蛋白质的间隔序列所隔开,这种mRNA叫做多顺反子mRNA。依笔者之见,原核生物多顺反子的存在,正好可视为是mRNA拼接成长链DNA的一个过渡阶段的证据。
另一个证据就是,在DNA聚合酶根据模板复制新的DNA链之前,必须依赖一段RNA引物。这一引物是引物酶辨认起始位点后解开一段DNA并按照5'到3'方向合成的RNA短链。之后,DNA聚合酶会通过磷酸二酯键的连接,添加与模板链配对的核苷酸,从而向引物链的3'端方向合成DNA。当然,最后RNA水解酶(RNase)会将RNA引物水解,另一些DNA聚合酶会生成DNA来填补这些缺口。
可以设想,如果不将m-RNA的遗传信息整合进一个统一的DNA中去的话,细胞分裂中如何能够将亲细胞准确地分配到两个子细胞中去则难以想象的!最近谢平(2015)提出,遗传密码子是生化系统的一部分,而生化系统的核心是ATP,正是ATP才建立起了核酸和蛋白质之间的联系(ATP中心假说)
当然,完善的遗传系统的建立绝非易事,超越了人类的想象,应该是细胞前体在数亿年的演化历程特别是无数次失败的分裂过程中才得以实现的。人们可能怀疑这种推论的真实性,但在如此宏大的地球上,在如此之小的细胞中,如果给予了10亿年的时日,一切偶然皆有可能成为必然,一切不可想象的事件皆有可能发生,只要有一个演化的方向性。
蛋白质、RNA和DNA三者在结构与功能的分化与完善,导致了一个完全独立的遗传系统的形成,而这又是通过细胞分裂维持生命形态相对稳定性的前提。只有一个真正可操作的遗传系统的出现,生命才从前细胞体时代迈进了细胞时代,才真正拉开了生物进化的序幕。
在生化机制上,细胞必须形成既有区别又有联系的一种结构体系,即一方面必须对信息进行准确地储存与复制,另一方面高效地实施生命构建,前者就是核酸体系,后者就是蛋白质体系。这两个体系在短时间尺度上相对独立,但不断相互作用,导致在长时间尺度上的协同演化。
RNA被写回DNA
美国托马斯·杰斐逊大学生物化学和分子生物学副教授Richard Pomerantz团队2021年6月刊登于《科学进展》的论文首次证明RNA也可以被写回DNA,这挑战了生物学的核心法则。